by George Wooten

Prepared for
Kettle Range Conservation Group
by George Wooten
Prepared for
Kettle Range Conservation Group
|
P.O. Box 150
Republic, WA 99166
509-775-2667
|
35 W. Main, Suite 220
Spokane, WA 99201
509-747-1663
|
We graciously acknowledge the support provided in the preparation of this report by the following organizations
The Kongsgaard-Goldman Foundation
The Bullitt Foundation
|
|
|
|
I. Executive SummaryII. BackgroundIII. Study MethodsIV. ResultsV. DiscussionVI. ReferencesAppendix A - Technical Procedures (not included here)Appendix B - Site descriptions (not included here)Appendix C - Photographs (not included here)
 |
At a time when many Northwest environmental organizations are primarily concerned with protection of forests, shrub-steppe lands of the Columbia Basin are quietly slipping away. A better understanding of the nature and function of shrub-steppe ecology is necessary to facilitate conservation efforts. What are shrub-steppe lands, how did they arise, how have they been altered, and why are they important? The answers to these questions give the necessary background for undertaking the challenge of protecting and preserving shrub-steppe lands for the benefit of future generations.
A. What are shrub-steppe lands?
Shrub-steppe lands of the interior Columbia Basin (also called the Columbia Plateau) comprise the northern tier of the semi-arid and essentially treeless biome that covers much of the interior west between British Columbia and Mexico. In Washington, the dominant vegetation is typically made up of various shrubs such as tall sagebrush (Artemisia tridentata), antelope bitterbrush (Purshia tridentata), rabbit brush (Chrysothamnus), winter fat (Atriplex) or other species of sagebrush (A. arbuscula, A. rigida). Grasses sometimes play a prominent role in these communities, including bluebunch wheatgrass (Pseudoroegneria spicata), Idaho fescue (Festuca idahoensis), needlegrasses (Stipa spp.) and Sandbergs bluegrass (Poa secunda). When grasses dominate, the lands are technically referred to as steppe. In addition, a diverse mixture of other plants and wildlife have developed within shrub-steppe habitats where they can take advantage of the plentiful sunlight and relative abundance of living space. Vegetation assemblages containing higher abundances of broadleaved forbs such as arrowleaf balsamroot (Balsamorhiza sagittata) are termed meadow-steppe. These types become prominent on the periphery of the Columbia Basin (Franklin and Dyrness, 1973), where the shrub-steppe grades into forested communities.
Much of our current understanding of Columbia Basin shrub-steppe ecology is due to Daubenmires (1970) application of a vegetation classification system based on the presence or absence of potential climax indicator species. His classification system grouped dominant vegetation types into a series of related plant associations, each arranged along a gradient of tolerance to limiting factors such as soil depth, precipitation or temperature. For instance, deep soils are a prerequisite for communities dominated by deep-rooted Idaho fescue (Festuca idahoensis), whereas shallow, jointed basalts limit growth to characteristic communities dominated by rock sagebrush (Artemisia rigida). Intermediate between these extremes plant species respond variably to form discreet communities that make up the various vegetation series.
Daubenmire determined that as a group, Washingtons shrub-steppe lands (literally, shrub-grasslands) are an artificial, descriptive aggregate, and that true steppe in the European sense, i.e., lands dominated by narrow-leaved grasses, were absent from his study area. This report focuses on Washingtons shrub-steppe, however much of the discussion applies throughout the interior west.
Taylor (1992) divided the shrub-steppe into six generalized ecological zones, each with a characteristic flora: (1) standard, a productive zone characterized by a lack of extremes, and which has been the most extensively lost to agriculture; (2) lithosols, with shallow basalt bedrock soils; (3) sand dunes; (4) talus; (5) meadows, moister areas that typically occupy depressions and; (6) saline soils, found in areas of low precipitation where mineral salts have accumulated on the soil surface.
In terms of both areal extent and ecological importance, one of the most significant plant communities with relevance to shrub-steppe lands is that of the lower ponderosa-pine (Pinus ponderosa) zone, also called the ponderosa pine-bunchgrass zone (Krajina, 1965; Brayshaw, 1965; Murphy, 1994; Arno and Hammerly, 1977). Ponderosa pine is important or prerequisite for a myriad of other species; in some habitats it is a keystone species which controls the development of the entire plant community. The transition zone between ponderosa pine and shrub-steppe is variable. In some cases the interface may be a gradual ecotone, in which case the boundary is somewhat arbitrary; in other cases the interface may form a mosaic (Hall, 1967). The lower pine zone presents a problem in landscape classification, not only because of the effect this has on area and ownership calculations, but also because the boundary between these communities is a dynamic one subject to change over time. The interface between ponderosa pine and shrub-steppe has moved significantly since white settlement began, partly in response to fire suppression and livestock grazing, but also due to long-term changes in climate and natural disturbances such as disease.
Unfortunately, no reliable map yet exists of Washingtons shrub-steppe that gives even a general level of detail at a meaningful scale. All existing maps incorporate assumptions that render accurate vegetation typing results as tenuous. For instance, it is difficult to tell healthy sagebrush communities from ones covering old plowed fields using just aerial photography. Therefore, the purpose of this report does not include classification of shrub-steppe vegetation beyond first approximations.
The purpose of this report is threefold: (1) to determine the extent of shrub-steppe lands as an aggregate group; (2) to serve as an initial step toward further conservation prioritization efforts which will require more detail to discriminate between habitat types and their relative degree of preservation; and (3) to determine as the ownership and preservation status of shrub-steppe lands.
It is important to consider shrub-steppe conservation regionally and over the long term. Knowledge about the location, extent and condition of shrub-steppe lands is indispensable for effective prioritization of preservation needs, as well as for the design of conservation programs which could conceivably involve land set-asides, conservation easements, or protective management designations.
B. How did Washingtons shrub-steppe arise?
The composition of shrub-steppe ecosystems is an important consideration in conservation of a representative subset of communities; this in turn is dependent on site conditions. The primary rock type within the Columbia Basin of Washington is the Columbia River Basalt Group, which originated from a series of basalt flows that erupted between 17 million and 6 million years ago from swarms of feeder dikes. The flows spread across a low-relief area of approximately 63,205 square miles and up to 4,000 meters (13,000 feet) thick, where they now abut mountains to the west, north and east. Following the eruptions, the basalts were faulted and folded to reliefs of up to 450 m (1,500 feet) (Campbell and Reidel, 1991, Tolan et. al., 1989, Griggs, 1978). The geological youth of these basalt rocks, their relative lack of breakdown and alteration, and their high degree of exposure at the surface, all provide a strong limitation on the species composition and physiognomy of vegetative communities.
Surrounding the Columbia basalts on three sides is a diversity of rock types, with the oldest types lying to the east (Griggs, 1978). To the west and north of the Columbia basalts, mountain ranges have arisen over the millennia through a complex of processes that include uplift, erosion, faulting, terrane accretion and volcanism (Stoffel et. al., 1991; Griggs, 1978, Tennyson and Cole, 1987). In contrast to the relatively shallow relief of the Columbia basalts, land at the margins of the flows exhibit greater variation in topography soils and microclimate; in response, landscapes along the edge of the Columbia Basin have developed with a high degree of biological diversity between and within plant communities. The biodiversity along the margin of the Columbia Basin is strongly manifest along tributary valleys and the Columbia River proper where shrub-steppe communities interfinger with montane and riparian communities that receive greater moisture and occupy different soils.
Species makeup and even entire communities have apparently changed radically since the end of Pleistocene Epoch and retreat of the ice sheet (Pielou, 1991). Evidence from pollen spectra measured in dated cores from the Okanogan Valley area indicate that sagebrush and graminoids were dominant while arboreal species were poorly represented during the first millennium after the ice retreated approximately 10,000 years ago (Mack et. al., 1989; Mack et. al., 1978; Alley, 1976). However extensive research indicates that these prehistoric plant assemblages have no modern analogue, even in the case of apparent sagebrush-steppe dominants (Mack and Bryant, 1974). The vegetative communities we know today began their formation about 6,700 years ago following the end of several thousand years of warm, dry climate, the hypsithermal (Pielou, 1991). From this point they may have attained their approximate present makeup by 3,000 years ago (Mack et. al., 1978; Alley, 1976; Hansen, 1940).
The Ice Age was also a time of massive meltwater flooding which radically altered the geology and vegetation patterns over the Columbia Basin. The most spectacular meltwater floods were the Spokane Floods, also known Missoula floods for the glacial lake of their origin, or as Bretz floods, after J Harlan Bretz, their discoverer. Bretz (1959) first discerned that the geology of Washingtons aptly named channeled scablands must have been due to flooding, the origin of which was due to periodic failures of ice dams holding back 2000 km2 of water in glacial Lake Missoula (Waitt, 1985).
The effect of the Spokane floods was profound. A network of meltwater channels was cut through bedrock hundreds of feet deep and as many miles long, reaching from the Idaho panhandle to the mouth of the Columbia and even into Oregon. The floods moved huge walls of rock and mud across the state, leaving behind a landscape of scoured bedrock, dry waterfalls, alluvial gravels the size of trucks, anomalous rock deposits left by rafted ice blocks, and ripple bars with 30 meter crests. Over the last 10,000 years, these flooded landscapes developed into unique plant communities, possibly even producing new species, for instance Hackelia hispida var. disjuncta (Hitchcock et. al., 1979; Gentry and Carr, 1976), which only occurs in large meltwater coulees.
In some areas the flood sediments have been locally reworked by wind to form dune sands or loess deposits (Reidel et. al., 1992). Another prominent soil feature which covers hundreds of square miles of central Washington soils are regularly spaced low mounds of fine soil atop a matrix of scoured basalt, known as biscuit-swale topography. This type of patterned ground has many competing hypotheses to explain its origin, for example intensive frost action associated with a periglacial climate (Kaatz, 1959).
These natural events combined to enable the formation of what we know of as the shrub-steppe biome of the Columbia Basin. Arno and Hammerly (1984) identified a number of factors that help maintain the treeless character of these areas: wind speed and duration; soils and geology; temperature; snow; precipitation; soil moisture; frozen ground; light intensity and biotic factors such as the lack of thermal protection from tree cover, and the lack of a seed bank for new tree establishment. Of these, the authors postulated the strongest determinants of tree exclusion to be precipitation, insolation (excessive heating) and cold.
Precipitation is the primary limiting factor for a number of species in the Columbia Basin. The height of the present Cascades Range presents a significant barrier to prevailing coastal moisture systems, leaving the east side of the mountains in a rain-shadow. The effect of the rain shadow is low precipitation and relative humidity in the Columbia Basin, with some areas receiving only 150 mm (6 inches) per year (Daubenmire, 1970). This severe aridity poses a serious constraint on species composition and growth habit within plant communities.
Taylor (1992) enumerated some of the adaptive traits which plants have evolved to survive in arid lands. Structural modifications possessed by some plants include a waxy leaf cuticle or leaf hairiness to resist solar and wind desiccation; small leaf size to reduce surface evaporation; succulence to provide water storage; moisture-obtaining structures such as modified roots; and modified anatomy to prevent dessication damage.
Many plants have modified lifestyles to grow preferentially during wet periods (Taylor, 1992). Grasses of the shrub-steppe employ a diverse range of specific strategies including tolerance of dehydration, efficient use of limited water, ability to withstand browsing, annual or ephemeral life cycles, and wind pollination. Tall sagebrush possesses both an evergreen set of leaves as well as a deciduous set which can take advantage of spring precipitation. Dense hairs on the leaves help prevent wind and solar desiccation. Sagebrush has both shallow roots to obtain water from summer thunderstorms as well as tap roots for reaching deep ground water.
The natural history of the Columbia Basin led to the development of many, diverse communities typically dominated by shrubs or grasses that are specialized for living in harsh, dry climates on a variety of soils. Many other species have adapted to these conditions, including invasive species which have fundamentally altered the function of the ecosystem, much to its and our detriment.
C. Why is it important to preserve shrub-steppe lands?
Shrub-steppe lands of the Columbia Basin once supported a nation of people. Today they help support a different nation. According to Turner et. al. (1980) Okanogan and other tribes traveled to the Columbia Basin to obtain many foods, utilitarian items and sacred materials indispensable to their cultures. Notable foods include serviceberries (Amelanchier alnifolia), bitterroot (Lewisia rediviva), camas (Camassia quamash) and Canbys lomatium (Lomatium canbyi). Wildlife and sources of tools (such as flint and obsidian) which are unique to certain areas in the Columbia Basin were also an integral part of their cultures. Today, domestic and ritual gathering of agricultural goods still continues as a viable cultural activity amongst native people living on the Colville Indian Reservation, however many traditional hunting, fishing and root-gathering grounds have been lost or compromised so profoundly that the cultural ties to the land are in danger of being permanently lost (Wooten, 2002, Turner et. al., 1980).
The resources of the shrub-steppe now provide amenities for a new nation. The most highly productive shrub-steppe soils are those of the Palouse loess, formed from deep, fertile, wind-blown soils which cover a large expanse of southeast Washington. But because of its fertility, the Palouse shrub-steppe has been completely transformed to wheat and other agriculture (Daubenmire, 1970), and its native peoples scattered to the winds. Of all the shrub-steppe habitats and cultures that have disappeared, the Palouse affords the most profound example of loss. But from the point of view of agricultural production, the area ranks as one of the most important in the U.S. Alarmingly, estimates of soil loss from the Palouse range as high as six inches a year. In the last hundred years, all of the original topsoil has been lost from about ten percent of the cropland, and from one-fourth to three-fourths of the original topsoil has been lost from another 60 percent of the cultivated cropland (Veseth, 1985).
Many species of plants and animals are unique to and dependent upon shrub-steppe habitats. Birds like the sage thrasher, mammals like the pygmy rabbit, and a host of other sagebrush-dependent species like the sage grouse and pygmy rabbit, which were once common, are now in decline (Ritter and Paige, 2000; Christensen, 2000; Washington Department of Wildlife, 1993). In order to stem the eventual loss of these species, enough habitat needs to be set aside for natural processes to continue.
Shrub-steppe lands offer boundless opportunities for recreation. Camping opportunities exist on many different ownerships and entire towns have economies based on recreation (Johnston, 2001). The Columbia Basin is a popular locality for the recreation sector including hunters, anglers, photographers, hikers, seekers of solitude and health advocates (Babcock and Carson, 2000).
The shrub-steppe has much to tell those who will listen. There is an awakening of interest in the shrub-steppe. Educational programs have been established that focus on the Columbia Basin ecosystem (Johnston, 2001; Ritter and Paige, 2000; Haynes et. al., 1996; Quigley et. al., 1996; USDA Forest Service, 1996). In the long term, education will be an indispensable aspect of shrub-steppe conservation, both for its contributions as well as its benefits.
D. How have Washingtons shrub-steppe lands been altered historically?
The natural disturbance regime and its mode of alteration occupies a central theme in discussions of conservation prioritization. Daubenmire (1970) provided important observations of human-caused changes which have affected natural succession in the Columbia Basin, including livestock grazing, European plant invasions and fire suppression. Any plan to preserve shrub-steppe habitat must incorporate consideration for the degree of altered function which can be tolerated.
Daubenmire cited historical accounts indicating that 200,000 livestock and feral horses were present in the Washington steppe by 1855. He observed that grazing accompanied the replacement of native species with annual grasses, and that invasion by alien species was transforming the Columbia Basin on a broad scale. He attributed a large share of the invasion damage to cheatgrass (Bromus tectorum) and Kentucky bluegrass (Poa pratensis), both of which have the potential for irreversible ecosystem alteration (Mack et. al., 2000; Wooten and Morrison, 1995). But Daubenmire found no evidence to support oft-made claims that sagebrush had increased its range during the settlement period.
Also lost from overgrazed soils are the fragile crusts of mosses, lichens and fungi known as cryptogamic, or cryptobiotic, crust, which Daubenmire found to be a significant element on almost all soil types. Soil crusts perform an irreplaceable ecosystem service by maintaining soil stability, retaining moisture, transferring nutrients, and maintaining shrub dominance (Bolton et. al., 1993; Perry et. al., 1989; Anderson et. al., 1982).
Daubenmire observed that over time, the suppression of fire in the shrub-steppe results in the gradual expansion of woody species. He cited anecdotal evidence from early explorers to indicate that wildfire was not omnipresent in the shrub-steppe, but a result of controlled burning by indigenous peoples.
The impact of indigenous peoples on the Columbia Basin was significant (Turner et. al., 1980). Each spring Sanpoil tribes (a tribe now living on the present-day Colville Indian Reservation) would move to the area south the Columbia River to dig roots over a 30-40 day period, over an area covering an estimated one million acres (Ray in Turner et. al., 1980, p. 147). Native activities in the Columbia Basin involved a number of ecosystem interactions, including digging, transportation, maintaining camps and the use of burning. O.C. Stewart observed, that all grasslands occurring on deep fertile soil are man-made, by peoples who periodically set fire to the grass and kept woody vegetation from growing (Daubenmire, 1970, p. 8).
The degree of degradation of shrub-steppe lands is an over-riding constraint in determining which lands deserve priority for preservation. In many cases, a misleading impression is given by figures which suggest large areas of Columbia Basin shrub-steppe remain, when in fact much of this land is so degraded as to be beyond restoration or ecological value. Part of this is due to the disparity between maps based on potential vegetation with that of the actual vegetation.
For instance, even though Quigley and Arbelbide (1997) concluded that the low sagebrush cover type is still as abundant as it was before 1900, many examples had severely altered successional pathways and the presence of invasive species. A slightly better understanding of the magnitude of loss is conveyed by the National Vegetation Classification, which lists 20% of Pacific Northwest dwarf shrub-steppe community as imperiled or critically imperiled (Anderson et. al., 1998). But it may be just as important to know what the condition is on the other 80% of dwarf shrub-steppe communities, because throughout the arid west, much of the land is in such poor ecological condition as to render ecosystem preservation a lost cause.
In many situations, the ability to reverse deleterious changes to shrub-steppe communities may not be practically feasible. McIver and Starr (2001) summarized cases of nearly insurmountable barriers to shrub-steppe restoration. These include loss of keystone animals prerequisite for seed dispersal or pollination (Longland, 1995, Whisenant, 1995), cases where native dominants or their seed sources are lacking (Laycock, 1991; West, 1999) and cases of altered fire regime (Mack et. al., 2000). The degradation process for shrub-steppe ecosystem components proceeds through a number of successive stages which are reversible only up to a limit. Beyond this limit, the community attains such highly altered ecological processes that passive restoration efforts involving mere removal of the causative agent would be ineffective.
Thus, measures of ecosystem value need to incorporate both negative attributes such as degraded condition along with positive attributes such as diversity and rarity.
III. Study Methods
A. Study Design Considerations
It is important to consider shrub-steppe conservation regionally and over the long term. Knowledge about the location, extent and condition of shrub-steppe lands is indispensable for effective conservation prioritization, as well as for the design of conservation programs which involve land acquisition, easements, or other protective management designations.
Examples of important questions which conservation prioritization might answer include the following:
The remaining objectives of this study are still underway. Toward that end, two important objectives that need the most work are (1) classification of shrub-steppe vegetation with higher reliability, resolution and detail; and (2) map overlays of shrub-steppe layers onto administrative ownerships.
B. Study preparations
Arcview 3.2 with Extensions was used as the main software platform for mapping analysis. View extents were assigned to be about 325 meters larger than the furthest boundary of any input file in any direction by use of a rectangular shape file, clipcov.shp. Because the offsite bounds of some of the digital maps extended reached far west into the Pacific ocean, Analysis Properties were set to match this shapefile as follows: cell size 82.02 ft; rows: 15,538; columns: 24,013; left extent: 941,767; right extent: 2,911,335; bottom extent: 81,550; top extent: 1,355,952.
Maps used were projected in the State Plane Coordinate System, Washington State Plane South Zone, Lambert Conformal Conic, Standard Parallel 45.833333, Longitude of Central Meridian 120.5, Latitude of Projection Origin (Reference Latitude) 45.333333, Geodetic Model NAD 1927, Clarke 1866 Ellipsoid. Planar distance units were in feet and cell size was 25 m (82.02 feet), except for the NHI-Historical map which was converted from a cell size of 1 km to one of 25m
A number of different sources for digital shrub-steppe maps were examined, and three of these were used for base maps in this study. These were: (1) GAP analysis maps for Washington, Version 5 digital land cover maps (Cassidy, 1997; also referred to here as lcv5 coverages); maps of shrub-steppe made by cooperatively by National Habitat Institute (NHI) and Washington Department of Fish and Wildlife (Kiilsgard , 1999, Kiilsgard and Barrett, 1999a and 1999b) and provided online by NHI as (2) NHI-Current and (3) NHI-Historical (presettlement, ca. 1850) maps.
The NHI and GAP maps were produced with different classification methods. The GAP map polygons were vector files made through photo-interpretation of major land cover and land use data within visually homogenous polygons derived from 1991 Thematic Mapper data, with a nominal minimum mapping unit of 100 hectares.
The NHI map are raster grid data derived from a rule-based classification scheme based on spectral and topographic attributes of the pixels. Nine Landsat Thematic Mapper scenes were used to classify cells of the NHI maps at a map scale of 1:100,000. All imagery contained less than 10% cloud cover and was acquired from May to October 1996.
NHI maps (as downloaded from the internet) are classified into 33 broad plant associations and landforms from an unsupervised maximum likelihood classifier algorithm, followed by vegetative typing and condition classfication using successive field verification in combination with ancillary data (such as topography) to refine the classification. Collateral data used included:
*2 Pinus ponderosa / steppe and Quercus garryi / steppe openings (Forest / steppe) 11 Festuca idahoensis/Rosa nutkana (Blue Mountains Steppe FEID/RONU) 12 Festuca idahoensis/Symphoricarpos albus (Palouse FEID/SYAL) 13 Artemisia tripartita/Festuca idahoensis (Three-tip Sage A. trip./FEID 14 Festuca idahoensis/Hieracium cynoglossoides (Klickitat Meadow Steppe FEID/Hier. cyno.) 15 Purshia tridentata/Festuca idahoensis (Bitterbrush PUTR/FEID) 16 Artemisia tridentata/Agropyron spicatum (Central Arid Steppe ARTR/AGSP) 17 Agropyron spicatum/Festuca idahoensis (Wheatgrass/Fescue AGSP/FEID) 18 Agropyron spicatum/Poa sandbergii (Canyon Grasslands AGSP/POSE) 19 Artemisia tridentata/Festuca idahoensis (Big Sage/Fescue ARTR/FEID)
A map layer of non-shrub-steppe cells was made by querying the GAP field Prim to create individual grids for developed, agricultural, deep water and wetlands. These grids were combined into a single theme and then clipped to match the extent of cells of the potential vegetation map ssgap5.
The determination the extent of shrub-steppe regional habitat types was performed by merging the grid nonss with ssgap5 and exporting the attribute values into a Microsoft Excel spreadsheet for calculations of area.
The determination of the extent of shrub-steppe overlap on major agency land owners in Washington was made using WSDOT (1995) coverages of Indian Reservations, Military sites, National Recreation Areas and National Forests. The files were first processed by reprojection. Next features of the agency coverage that intersected the selected shrub-steppe polygons were selected and with the selected themes active, the Arcview Summarize Zones command was used to determine the overlap of those polyons with the grid cells in the shrub-steppe grid ssgap5. Results were exported to a Microsoft Excel spreadsheet with the following results (data is rounded to the nearest mile unless it is less than one square mile or zero).
IV. Results
Consideration of error sources
Errors considered included input mapping errors, projection errors, round-off errors, use of data at different scales and misregistration of different layers. The latter two were corrected by assigning a common analysis grid for cell alignment. Round-off errors were minimized by using a minimum of 6 significant figures for all calculations. Mapping errors were acknowledged in the source data. Projection errors were assessed as follows.
The extent and accuracy of the grid file unit sizes were checked by summing all cells for the entire state of Washington, which is given as 71,303 sq miles.
The extent of all Washington counties, using the 25 m cells of Washington State Department of Transportation county map, co_grid (WSDOT, 1995), yielded 295,553,779 cells or 71,320 sq mi (184,720 sq km), an error of about 0.02%. Considering that the map involves projection across two state plane zones, this is a respectable figure. However, this figure includes territorial waters, coastal bays and estuaries, which extend miles out into the waters of Puget Sound and the Pacific Ocean. It was desirable to know the land surface of Washington state, exclusive of coastal waters. When coastal waters were selected out of the GAP 25 m grids, it gave the extent of Washingtons land surface and inland waters as 67,653 sq mi, or about 5% lower. The NHI-Current maps yield a similar figure for Washingtons land-only extent as 67,640 sq mi.
The lower figures are more representative of the actual, as opposed to territorial, land area of Washington, although they still includes the territorial waters of the Columbia and Snake River boundaries with Oregon. This analysis project used the mean of the two lower figures as a best estimate of the actual land area of Washington, or 67,647 sq mi (175,204 sq km).
Extent of shrub-steppe lands
Several different estimates of past and current shrub-steppe extent were made. The NHI-Historical maps indicated that 23,711 sq mi (61,411 sq km) or approximately 35% of the total land area of Washington, was originally shrub-steppe. The hybrid GAP-NHI potential vegetation layer ssgap5 indicated the original shrub-steppe extent to be 24,437 sq mi, (approximately 36% of Washington) which is about 3% (or 726 sq mi) of the estimate made using the NHI-Historical map.
The NHI-Current maps indicated that 12,701 sq mi (32,897 sq km, or about 53.6%) of Washington shrub-steppe remains from the original extent given by the NHI-Historical map. If the percentage of shrub-steppe cells on the NHI-Current maps included only those that were previously classified as shrub-steppe on the NHI-Historical map, then the area of existing shrub-steppe would be 41,167,724 cells (9,934 sq mi), or 41.9% of the original. And if the percentage of shrub-steppe cells on the NHI-Current maps included only those that were classified as shrub-steppe on the grid of potential shrub-steppe vegetation, ssgap5, then 11,173 sq mi or 45.7% of original shrub-steppe remain.
Subtraction of GAP agricultural, developed, water
and wetland areas from the hybrid GAP-NHI map ssgap5 indicates that
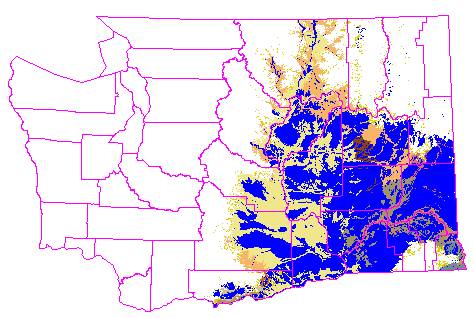
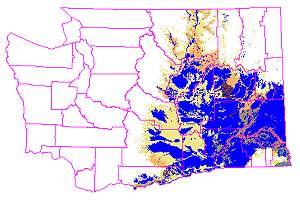
11,315 sq mi (46.3%) of the GAP potential shrub-steppe vegetation remains
unconverted, as illustrated in this image, which shows shrub-steppe lands
that have been converted to agriculture and development in blue, and remaining
shrub-steppe vegetation in light tan shades.
 |
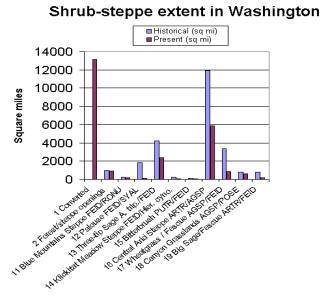
The area of each category of converted shrub-steppe lands (non_ss) which were subtracted from the map of GAP potential shrub-steppe vegetation (ssgap5) is given below.
|
|
|
|
|
Developed
|
1,013,639
|
245
|
|
Agricultural
|
50,604,723
|
12,211
|
|
Water
|
1,847,034
|
446
|
|
Wetlands
|
912,579
|
220
|
|
Total
|
13,122
|
|
|
Historic (sq mi) |
Existing (sq mi) |
|
1
Converted
|
0
|
13,122
|
|
2
Shrub-steppe forest openings
|
973
|
920
|
|
11
Blue Mountains Steppe FEID/RONU
|
227
|
157
|
|
12
Palouse FEID/SYAL
|
1,805
|
151
|
|
13
Three-tip Sage A. tripartita/FEID
|
4,197
|
2,410
|
|
14
Klickitat Meadow Steppe FEID/Hieracium cynoglossoides
|
243
|
89
|
|
15
Bitterbrush PUTR/FEID
|
94
|
47
|
|
16
Central Arid Steppe ARTR/AGSP
|
11,927
|
5,874
|
|
17
Wheatgrass/Fescue AGSP/FEID
|
3,363
|
871
|
|
18
Canyon Grasslands AGSP/POSE
|
812
|
602
|
|
19
Big Sage/Fescue ARTR/FEID
|
794
|
194
|
|
TOTAL
|
24,435
|
24,437
|
 |
|
|
Historical (sq mi) |
Existing (sq mi) |
|
|
Colville
Indian Reservation
|
765
|
93
|
12.2%
|
|
Spokane
Indian Reservation
|
18
|
0.07
|
0.39%
|
|
Yakama
Indian Reservation
|
1,036
|
352
|
34.0%
|
|
Fairchild
Air Force Base
|
1.01
|
0.47
|
46.5%
|
|
Military
Yakima Training Center
|
503
|
2.83
|
0.56%
|
|
Department
Of Energy Hanford
|
585
|
34
|
5.8%
|
|
Coulee
Dam NRA
|
39
|
27
|
69.2%
|
|
Lake
Chelan NRA
|
0.27
|
0
|
0%
|
|
Colville
NF
|
15
|
1.09
|
7.3%
|
|
Okanogan
NF
|
135
|
1.33
|
1.0%
|
|
Non-NF
Inholding
|
3.33
|
0.01
|
0.3%
|
|
Kaniksu
NF
|
0.0029
|
0
|
0%
|
|
Wenatchee
NF
|
153
|
0.25
|
0.16%
|
|
Umatilla
NF
|
38
|
0
|
0%
|
|
TOTAL
|
3,291
|
512
|
15.6%
|
 |
V. Discussion
Assumptions
The potential vegetation map ssgap5 forms the basis for calculations of shrub-steppe extent. It is extremely important to be mindful of the assumptions embodied in the potential vegetation map ssgap5. It is a hybrid combination of GAP potential shrub-steppe vegetation types with GAP potential ponderosa pine and oak forest cells which overlapped existing NHI shrub-steppe types.
The preference was to use GAP map layers since these were manually digitized and the error is known to a greater degree. However, the GAP polygons tended to be overly inclusive of minor areas that did not match the overall attributes of the polygon, and furthermore the GAP input data is about five years older than the NHI data (1991 versus 1996). For the purpose of this project, errors of commission were judged to be preferable to errors of omission, in both the determination of shrub-steppe as well as agricultural land extent. It was desirable to have a potential shrub-steppe map that was slightly over-inclusive in order to capture information about rare vegetation associations at the boundary of the shrub-steppe, as well as to account the for lands encompassing the original extent of shrub-steppe, about which the boundaries are unknown and inprecise. It was desirable to have an agricultural layer that was over-inclusive because both the GAP and NHI input maps were conservative in their estimates of agricultural conversion. Closer examination of the shrub-steppe maps revealed that fallow lands were sometimes misclassified as shrub-steppe. Also, most of the existing shrub-steppe lands in Washington have been altered by livestock grazing, which typically was not counted as agriculture.
The large number of assumptions went into the calculations of past and present shrub-steppe extent mean that the figure of 11,315 square miles or 46.3% of remaining shrub-steppe is too optimistic. At this point, other factors that would lower this figure include the following: (1) the input data is ten years old; (2) the historic extent of the shrub-steppe did not include small peripheral areas; (3) agricultural areas were defined to be 100% converted--other lands counted as shrub-steppe were severely degraded rangelands, fallow fields, weed infestations and agricultural areas with less than 100% conversion. Similarly, one could enumerate unaccounted factors that raise amount of existing shrub-steppe, such as failure to include wet areas in the historic shrub-steppe, while subtracting these out from the present shrub-steppe, however these factors account for a much smaller margin of error than those that lower the amount of existing shrub-steppe. To account for these factors, further research in vegetation typing and condition mapping needs to be accomplished.
Shrub-steppe layer development
The addition of the layer, Shrub-steppe openings in ponderosa pine and oak forests, to the GAP shrub-steppe grid layer was done because an examination of the GAP ponderosa pine and oak layers determined that significant areas of shrub-steppe openings within the two lower montane forest types could be reliably identified by the NHI shrub-steppe classification. This is also reasonable in an ecological sense, since ponderosa pine and Garry oak forests generally intergrade with shrub-steppe lands as an ecotone.
Discrepancies in interpreting the data
The GAP agricultural layer was chosen over the NHI agriculture layer for use in this project mainly for expediency, as it was already contiguous with and not overlapping the other non-shrub-steppe categories. Another reason for using the GAP agricultural layer was that in areas where the two maps differed, the NHI agricultural layer tended to contain many small groups of cells that were apparently artifacts of the classification. Further refinement of the agricultural layer is recommended, perhaps by refinement of the NHI layer or development of a layer containing attributes for relative degree of alteration.
Even though the NHI maps were used to a lesser extent than the GAP maps, the different estimates of shrub-steppe conversion using those maps gives a good example of the problems associated with mapping errors.
The percentage of 53.6% (12,701 sq mi) of remaining shrub-steppe made by subtracting the NHI-Current from the NHI-Historic maps erred too high because the NHI-Historical map periphery does not extend as far as the NHI-Current map does, leading to a large number of shrub-steppe cells that would seem to be increasing beyond their original boundary.
If the percentage of shrub-steppe cells on the NHI-Current maps included only those that were previously classified as shrub-steppe on the NHI-Historical map, then the percentage of existing shrub-steppe would be 9,934 sq mi, or 41.9%, of the original. This lower figure is probably closest to the actual percentage of remaining shrub-steppe, however it errs by not considering the cells outside the original periphery of historical shrub-steppe.
If the percentage of shrub-steppe cells on the NHI-Current maps is calculated only from those that overlap the GAP grid ssgap5 of potential shrub-steppe vegetation, then 11,173 sq mi, or 45.7%, of original shrub-steppe remain. This errs in that it uses circular reasoning. The potential shrub-steppe layer attribute for openings in the ponderosa pine and oak forests was derived from the same source for both the potential and existing maps of shrub-steppe. While it is true that this layer (ssnonfor) was derived from a combination of two inputs so as to be as accurate as possible, it renders calculation of the loss of historical shrub steppe meaningless for those cells. Nevertheless, this figure of existing shrub-steppe extent is probably more accurate because of the circular reasoning--it is the calculation of conversion that is flawed, and this only for the non-forest openings attribute.
The results of mapping the extent of regional shrub-steppe habitat types does not contribute much to the understanding of shrub-steppe ecology. These habitat types are essentially just areas of regional similarity, within which many different true vegetative associations exist. To be useful, the maps need to provide habitat typing at a higher level of detail. Even the six generalized habitat types given in the background of this report would provide a better estimate of the relative extent of different types of vegetation since those use physiognomic descriptors rather than regional ones.
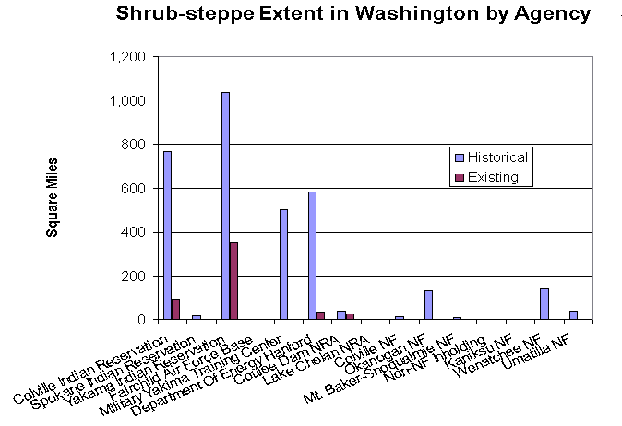
The results of the analysis of administrative ownership of shrub-steppe lands was informative in that it revealed that the four groups of agencies analyzed are doing a poorer job of protecting shrub-steppe habitat than the other ownerships that werent analyzed (including private lands). The overall percentage of shrub-steppe conservation on these lands was a mere 15.6%, less than half of the percentage of the norm. If true, this reflects badly on the role of these agencies as land stewards. Only the Coulee Dam RNA, at 69.2% shrub-steppe remaining, was significantly above the norm. It was unexpected to find that the large Colville and Yakama Indian Reservations, with their vested interested in native cultural use of shrub-steppe lands, were below the norm in shrub-steppe preservation.
These findings need corroboration, and if true, some adjustments to management or management goals may be in order. In any case, the findings should be a stimulus toward further elucidation of the ownership and management of shrub-steppe lands and which shrub-steppe lands are currently receiving protection.
Conclusion
From the results of this analysis, an overly optimistic portrayal is given that roughly half of the original shrub-steppe lands of Washington remain intact. This is misleading because many of those lands are in such poor condition as to be without much ecological value. Examination of field sites on the existing shrub-steppe layer revealed that some shrub-steppe lands have become degraded beyond conservation value.
Bolton, H. Jr., J.L. Smith, S.O. Link (1993). Soil microbial biomass and activity of a disturbed and undisturbed shrub-steppe ecosystem. Soil Biol. Biochem. 25:545-552.
Brayshaw, T.C. (1965). The dry forest of southern British Columbia. In K.Greenridge (ed.), Essays in plant geography and ecology, pp. 167-184. N.S. Museum, Halifax, N.S., Can.
Bretz, J (1959). Washington's channeled scabland. Washington Div. Mines and Geol. Bull. 45.
Campbell, Newell, and Stephen Reidel (1991). Geologic Guide for Star Routes 240 and 243 in South-Central Washington, Washington Geology, 19:3-17.
Cassidy, K.M. (1997). Washington Gap Project 1991 Land Cover for Washington State, Version 5; Published by Washington Cooperative Fish and Wildlife Research Unit, University of Washington, Seattle (http://www.wa.gov/wdfw/wlm/gap/landcov.htm).
Christensen, J. (2000). Fire and cheatgrass conspire to create a weedy wasteland. High Country News, 32(10), May 22, 2000.
Daubenmire, Rexford (1970). Steppe Vegetation of Washington. Wash. Agr. Expt. Sta. Tech. Bull. 62, Washington State University, Pullman, WA. 131 p.
Franklin, Jerry F., C.T. Dyrness (1973). Natural Vegetation of Oregon and Washington. USDA-FS PNW For. and Range Expt. Sta., repr. Oregon State Univ. Press, Corvallis, OR. 452 p.
Gentry, Johnnie and R. Carr (1976). A revision of the genus Hackelia (Boraginaceae) in North America, north of Mexico. Memoirs of the New York Botanical Garden 26(1):178-181.
Griggs, A.B. (1978). Columbia Basin. Pp. 22-27 in Livingston, Vaughn, Jr., 1978, Geology of Washington, State of Washington Department of Natural Resources, Division of Geology and Earth Resources, Reprint 12, prepared in cooperation of U.S. Geological Survey, reprinted from a report prepared for the U.S. Senate Committee on Interior and Insular Affairs in 1966, Mineral and Water Resources of Washington.
Hall, Fred (1967). Vegetation-soil relations as a basis for resource management on the Ochoco National Forest of central Oregon. Unpublished M.S. thesis on file at Oregon State Univ., Corvallis.
Hartnett, D.C., G.W.T. Wilson (1999). Mycorrhizae influence plant community structure and diversity in a tallgrass prairie. Ecology 80(4): 1187-1195.
Haynes, R.W., R.T. Graham, T.M. Quigley, Tech. Eds. (1996). A framework for ecosystem management in the Interior Columbia Basin and portions of the Klamath and Great Basins. USDA-FS, PNW Region, GTR-PNW-GTR-374, Portland, OR.
Hitchcock, C. Leo, A. Cronquist, M. Ownbey, J.W. Thompson, illus. J.R. Janish (1969). Vascular Plants of the Pacific Northwest. Univ. of Wash. Press, Seattle, WA Vols. 1-5.
Hooge, Philip (1996). Spatial Tools Arcview Extension. USGS - BRD, Alaska Science Center - Biological Science Office Glacier Bay Field Station, (http://www.absc.usgs.gov/glba/gistools/spatialtools_doc.htm).
Johnston, Greg (2001). Painted desert; Seattle Post-Intelligencer, Getaways, pp. 12-16, May17, 2001.
Kaatz, Martin (1959). Patterned ground in central Washington: a preliminary report. Northwest Sci. 33:145-156.
Kiilsgard, Chris, Northwest Habitat Institute (1999). Washington Wildlife Habitat Types: Mapping and Classification of Landscape-Level Habitats, Final Report Submitted to Washington Department of Fish and Wildlife: Habitat Program. Northwest Habitat Institute, P.O. Box 855, Corvallis, OR, 97339.
Kiilsgard, Chris, and Barrett, Charley, Northwest Habitat Institute (1999a). Washington Current habitat types (wa_curr). Maps were made by Washington Department of Fish and Wildlife and provided online by Northwest Habitat Institute, P.O. Box 855, Corvallis, OR, 97339 (http://www.nwhi.org/nhi/).
Kiilsgard, Chris, and Barrett, Charley, Northwest Habitat Institute (1999b). Washington Historic circa 1850 habitat types (wa_hist). Maps were made by Washington Department of Fish and Wildlife and provided online by Northwest Habitat Institute, P.O. Box 855, Corvallis, OR, 97339 (http://www.nwhi.org/nhi/).
Krajina (1965). Biogeoclimatic zones and classification of British Columbia. In V.J. Krajina (ed.), Ecology of Western North America. 2:1-17; UBC Dept of Botany.
Laycock, W.A. (1995). New perspectives on ecological condition of rangelands: can state and transition or other models better define condition and diversity?. In: Montes, L., Olivia, G.E. (Eds.), Proceedings of the International Workshop on Plant Genetic Resources, Desertification and Sustainability. INTA-EEAA, Rio Gallegos, Argentina.
Lincoln, R.J., G.A. Boxshall, P.F. Clark (1990). A Dictionary of Ecology, Evolution and Systematics. Cambridge Univ. Press, New York, NY. 298 p.
Longland, W.S. (1995). Desert rodents in disturbed shrub communities and their effects on plant recruitment. In: Montes, L., Olivia, G.E. (Eds.), Proceedings of the International Workshop on Plant Genetic Resources, Desertification and Sustainability. In: Roundy, B.A., McArther, E.D., Haley, J.S., Man, D.K. (Comps.), Proceedings of the Wildland Shrub and Arid Land Restoration Symposium, Las Vegas, NV, Oct 19-21. USDA-FS report INT-GTR-315, Intermountain Research Station, Ogden, UT.
Mack, R., D. Simberloff, W.M. Lonsdale, H. Evans, M. Clout, F. Bazzaz (2000). Biotic invasions: causes, epidemiology, global consequences and control. Issues in Ecology. No. 5, Ecological Society of America, Washington D.C.
Mack, Richard N., N.W. Rutter, S. Valastro (1979). Holocene vegetation history of the Okanogan Valley, Washington. Quat. Res. 12:212-225.
Mack, Richard N., V.M. Bryant Jr. (1974). Modern pollen spectra from the Columbia Basin, Washington. Northwest Sci. 48:183-194.
Mack, Richard N., N.W. Rutter, S. Valastro (1978). Late quaternary pollen record from the Sanpoil River, Washington. Can. J. Bot. 56:1642-1650.
McIver, James, Lynn Starr (2001). Restoration of degraded lands in the interior Columbia River basin: passive vs. active approaches. Forest Ecology and Management 153:15-28.
Murphy, Alexandra, illus. R. Petty (1994). Graced by Pines - The Ponderosa Pine in the American West. Mountain Press Publishing, Missoula, MT.
Perry, David A., M.P. Amaranthus, J.G. Borchers, S.L. Borchers, R.E. Brainerd (1989). Bootstrapping in ecosystems. Bioscience 39:230-237.
Pielou, E.C. (1991). After the Ice Age. The Return of Life to Glaciated North America. Univ. of Chicago Press, Chicago.
Quigley, T. M., and S. J. Arbelbide, technical editors (1997). An assessment of ecosystem components in the interior Columbia basin and portions of the Klamath and Great Basins. Volume 2. U.S. Forest Service General Technical Report PNW-GTR-405.
Quigley, T.M., R.W. Haynes, R.T. Graham, T. Russell, eds. (1996). Integrated Scientific Assessment for Ecosystem Management in the Interior Columbia Basin and Portions of the Klamath and Great Basins. USDA-FS PNW-GTR-382, Portland, OR.
Reidel, S. P., K.A. Lindsay, and K. R. Fecht (1992). Field Trip Guide to the Hanford Site. WHC-MR-0391, Westinghouse Hanford Company. Richland, Washington
Ritter, Sharon, and Christine Paige (2000). Keeping birds in the sagebrush sea. Joslyn and Morris, Boise, ID (available from the Wenatchee BLM with a video titled, The Vanishing Shrub-Steppe.
Stoffel, K.L., compiler (1990). Geologic map of the Republic 1:100,000 quadrangle, Washington: Washington Diuvision of Geology and Earth Resources Open File Report 90-10, 62 p., 1 pl.
Taylor, Ronald J (1992). Sagebrush Country - A Wildflower Sanctuary. Mountain Press Publishing, Missoula, MT.
Tennyson, Marilyn, and Mark Cole (1987). Upper Mesozoic Methow-Pasayten sequence, Northeastern Cascade Range, Washington and British Columbia. Washington Division of Geology and Earth Resources Bulletin 77:73-84.
Tolan, T.l., Reidel, S.P., Beeson, M.H., Anderson, J.L., Fecht, K.R., Swanson, D.A. (1989). Revisions to the estimates of the areal extent and volume of the Columbia River Basalt Group. In, Reidel, S.P., Hooper, P.R., eds., Volcanism and tectonism in the Columbia River flood-basalt province. Geological Society of America Special Paper 239, p. 1-20.
Turner, N.J., R. Bouchard, D. Kennedy (1980). Ethnobotany of the Okanagan-Colville Indians of British Columbia and Washington. Occasional Papers of the British Columbia Provincial Museum, No. 21, B. C. Provincial Museum, Victoria, B. C.
Schäuble, Holger, University of Tübingen (Institute of Geography), Germany (1998). Grid Enhancement Extension (GRID Tools) (http://www.uni-tuebingen.de/uni/egi/studium/projekte/schaeuble/sites/grid.htm)
US EPA (1998). Guidelines for Ecological Risk Assessment, Document EPA/630/R-95/002F, Federal Register 63(93):26846-26924, May 14, 1998 (http://www.epa.gov/ncea/raf/pdfs/ecotxtbx.pdf).
USDA Forest Service (1996). Status of the interior Columbia basin: summary of scientific findings. USDA-FS PNW-GTR-385, Portland, OR.
Veseth, Roger (1985). Erosion impacts on the Palouse misunderstood. PNW Conservation Tillage Handbook Series; WSU Direct Seed Tillage Handbook (http://pnwsteep.wsu.edu/tillagehandbook/chapter1/010185.htm).
Waitt R.B. (1985). Case for periodic colossal jökulhlaups from Pleistocene Lake Missoula; Geol. Soc. Am. Bulletin, v. 96, pp. 1271-1286.
Washington Deptartment of Wildlife (1993). Pygmy rabbit (Brachylagus idahoensis) in Washington. Washington Deptartment of Wildlife, 600 Capitol Way N., Olympia, WA.
West, N.E. (1999). Managing for biodiversity of rangelands. Pp. 101-126 In: Collins, W.W., Qualset, C.O. (Eds.), Biodiversity in Agroecosystems. CRC Press, Boca Raton, FL.
Whisenant, S.G. (1995). Landscape dynamics and arid land restoration. In: Roundy, B.A., McArther, E.D., Haley, J.S., Man, D.K. (Comps.), Proceedings of the Wildland Shrub and Arid Land Restoration Symposium, Las Vegas, NV, Oct 19-21. USDA-FS report INT-GTR-315, Intermountain Research Station, Ogden, UT.
Wooten, George (2002). (personal conversation with tribal members from Nespelem during the root-digging season).
Wooten, G.F., P.H. Morrison (1995). Biological invasions of alien plants in the interior Columbia River Basin (http://www.okanogan1.com/references/aliens.htm). In: Cara Nelson, ed., Key Elements for Ecological Planning: Management Principles, Recommendations, and Guidelines for Federal Lands East of the Cascade Crest in Oregon and Washington, Columbia River Bioregion Campaign, Science Working Group, Walla Walla, WA.
WSDOT (Washington State Department of Transportation) (1995). Counties of Washington State (http://www.wsdot.wa.gov/mapsdata/geodatacatalog/default.htm).

A summer thunderstorm brews over the Colville Indian Reservation.